 切换视图
切换视图

- 模式产地: 中国:湖北:宜昌
- 模式标本: 全模标本 ( 2号 ) , 选模标本 ( BMNH 1947.2.3.86 ) 保存于BMNH ( The Natural History Museum, London, UK )
- 体肥壮;雄蛙胸、腹部满布大小黑刺疣。
- 体长 : 雄蛙体长78–100mm,雌蛙体长89–111mm,大者达124mm
- 体型 : 体型甚肥硕
- 头 : 头宽大于头长
- 吻 : 吻端圆,略突出于下唇
- 吻棱 : 吻棱不显
- 鼻孔 : 鼻孔位于吻眼之间
- 鼻间距vs吻眼距vs上眼睑宽 : 眼间距和鼻间距几乎相等,小于上眼睑宽
- 瞳孔 : 瞳孔呈菱形
- 鼓膜、鼓环 : 鼓膜不显
- 颞褶 : 颞褶甚厚
- 舌 : 舌椭圆形,后端有缺刻
- 犁骨齿 : 犁骨齿短,呈“\ /”
- 声囊 : 具单咽下内声囊;声囊孔大,长裂状
- 背侧褶 : 无背侧褶
- 前肢 : 前肢粗壮
- 前臂 : 前臂及手长不到体长之半
- 指、趾 : 指略短
- 指关节下瘤 : 关节下瘤甚明显
- 内、外掌突 : 内掌突大卵圆形,外掌突窄长
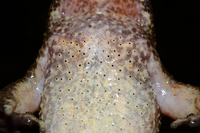
- 第二性征 : 雄蛙前臂极粗壮,内侧3指有锥状黑刺;胸腹部满布大小刺疣,疣上有刺一枚
- 雄性线 : 背侧有紫色雄性线
- 后肢 : 后肢肥壮
- 后肢贴体前伸 : 前伸贴体时胫跗关节达眼部
- 左右跟部是否相遇 : 左右跟部仅相遇
- 脚长vs胫长vs体长 : 胫长略超过体长之半
- 指、趾端 : 指、趾端圆球状,无沟
- 指、趾缘膜 : 第二指两侧及第三指内侧具缘膜;第一、五趾游离侧缘膜达蹠基部
- 指、趾式 : 指式:3>1≈4>2
- 指、趾蹼式 : 趾间几乎全蹼,第四、第五蹠间蹼超过蹠长之半
- 趾关节下瘤 : 关节下瘤明显
- 蹠趾腹面疣 : 有跗褶
- 内、外蹠突 : 内蹠突窄长,无外蹠突
- 皮肤、体色 : 体背面皮肤粗糙,背部有纵行的长形或圆形疣和小刺疣;眼后有一横肤沟;四肢背面疣少,长形肤棱明显;体色有变异,背面多为黄棕色或深褐色,两眼间常有一黑横纹,有的背部具不规则的黑斑;四肢背面有黑横纹或不明显;体和四肢腹面肉色,咽喉部有棕色斑,下颌缘更明显
- 卵 : 卵径4mm左右,动物极灰棕色,植物极乳黄色。卵胶膜3层,卵间以外层胶膜彼此贴连,以数十粒连成串状似葡萄,也有个别卵为单粒状
- 蝌蚪 : 第36–38期蝌蚪全长平均52mm,头体长19mm左右,尾长约为头体长的179%;体背面黄棕色或褐黑色,尾部色较浅有灰色斑点,体尾交界处有一黑横斑;头体背腹扁,尾肌发达,尾鳍平直,尾末端钝尖;唇齿式多为Ⅰ:4+4/1+1:Ⅱ。上唇无乳突,下唇乳突两排,口角有副突
- 生境: 该蛙生活于海拔300–1900m山区的流溪或其附近的水塘中。
- 习性: 白天隐匿于溪底的石块下、溪边大石缝或瀑布下的石洞内;晚间出外,蹲于石块上或伏于水边,夏季常发出“梆、梆、梆”的洪亮鸣声。该蛙以捕食昆虫为主,其有益系数为68.3%。卵多产于流溪瀑布下水坑内,卵群成串,似葡萄状,一般黏附在水中石块下、倒木或枯枝上。蝌蚪生活于流溪水坑内;冬季多潜伏在水底石下或落叶层下越冬。
- 繁殖季节: 5月,6月,7月,8月

























































- 陕西;山西;甘肃;四川;重庆;云南(昭通、陆良、大关、河口、巧家、屏边、威信、绥江、彝良、西畴、屏边、马关);贵州习水国家级自然保护区;湖北;江西;湖南;广西
- 同物异名: Rana Boulengeri, Rana tibetana, Rana robertingeri(合江棘蛙), Nanorana robertingeri, Nanorana boulengeri, Quasipaa robertingeri, Quasipaa tibetana
- 俗名: 石棒、石坎
-
基于形态学研究, 吴贯夫和赵尔宓 (1995) 将采自四川省合江县天堂坝村的棘蛙个体描述为合江棘蛙。费梁等 (2012) 也收录合江棘蛙为有效物种 (合江棘蛙: 466-467页; 棘腹蛙: 460-461页)。Che et al. (2009) 基于分子系统学研究提示合江棘蛙可能不是一个有效的物种: 线粒体基因片段 (12S, 16S rRNA) 分析 (Fig. 5) 以及线粒体与核基因片段 (12S, 16S rRNA,Rhod,Tyr) 的联合分析 (Fig. 6) 均显示合江棘蛙 (YNU-HU20025105, YNU-HU20025106) 位于棘腹蛙支系的基部位置, 但核基因数据 (Fig. 4) 显示二者聚在一起, 无法区分。Yan et al. (2013) 利用线粒体基因 (Cytb) 和核基因 (GCG) 对棘腹蛙进行了种群系统地理学研究, 样品覆盖了该物种所有分布区域, 同时包含了合江棘蛙地模样品(见该文Materials and Methods)。线粒体基因数据分析显示, 棘腹蛙由4个支系 (A-D) (Fig. 2) 组成, 支系A (样品来自四川合江、叙永和贵州赤水) 位于基部位置, 与其他支系B-D之间的分化程度较大, 此外, 该研究也发现来自四川合江的棘蛙分属两个线粒体支系A和C1 (采集点10, Fig. 1; Table S1); 抽取部分样品构建的核基因等位基因树和中介网络图均识别出两个高支持率的进化支:AI和AII (Fig. 3), 和线粒体4个支系划分并不一致, 其中AI包括了线粒体支系A的所有个体和支系B-D的部分个体。来自合江地区的分属线粒体支系A (KIZSC0709008) 和C1 (KIZSC0709006) 的棘蛙个体位于核基因进化支AI中 (Fig. 3, Table S1)。这一结果提示, 合江地区的棘蛙虽然出现线粒体基因片段的高度分化, 但核基因数据支持合江地区的棘蛙与棘腹蛙不同种群之间存在基因交流。Yan et al. (2021) 在上述以及Che et al. (2010) 的研究基础上, 基于线粒体 (Cytb) 和核基因 (Rag1, Rag2, Rhod, Tyr) 片段构建了迄今棘胸蛙属物种最为全面和清楚的系统发育关系, 其中基于四个核基因片段的联合数据非常有力的恢复了研究中所用的棘胸蛙属各个物种的单系, 并较好的解决了它们的系统发育关系 (Fig. 2a)。该研究中, 支系D包括两个物种, 一个是未描述物种Q. sp2, 另一个是棘腹蛙, 这两个物种的单系性得到了线粒体 (Fig. 2b, Fig. S1) 以及核基因 (Fig. 2a) 的共同支持。在棘腹蛙种群中, 贵州赤水的其中两个个体单独形成一个线粒体支系 (KIZ07984、KIZ07980对应Yan et al., 2013中线粒体支系A), 另一个体 (KIZ07978) 与四川合江 (KIZSC0709003, KIZSC0709006) 及其它棘腹蛙种群形成另一独立的线粒体支系 (Fig. S1); 但是, 与线粒体结果不同, 核基因构树显示棘腹蛙种群无明显结构分化, 来自贵州赤水属于不同线粒体基因支系的棘蛙 (KIZ07980, KIZ07978) 与棘腹蛙其它种群聚在一起, 分化不明显 (Fig. 2a)。综上, 尽管来自四川合江和贵州赤水等地的棘蛙存在明显的线粒体支系分化, 但基于核基因的研究显示它们与棘蝮蛙其它种群分化不显著, 存在共享核基因单倍型, 提示具有基因交流。Wang et al. (2021) 扩大采样地点, 对四川、贵州和重庆地区的合江棘蛙和棘腹蛙开展了研究。综合GENEBANK数据, 该研究基于线粒体COI基因片段分析发现合江棘蛙和棘腹蛙分别形成独立的支系 (Fig. 1; Q. robertingeri, Q. boulengeri), 从而建议恢复合江棘蛙有效性。鉴于上述分子系统学的研究, 如果仅看“合江棘蛙是否有效”这一问题, 不同研究的结论显然是不一致的。为了解决这些疑问, 我们重新分析了数据。首先, 我们发现Wang et al. (2021) 的研究未对Yan et al. (2013) 的工作进行引用和相应的分析及讨论, 忽略了核基因数据中反映出来的基因交流现象。其次, 尽管不同研究使用了不同的基因片段, 但仔细对比不同研究发现, 合江棘蛙对应的线粒体独特支系的系统发育位置在四篇工作中是一致的: Che et al., 2009 (P. robertingeri), Yan et al., 2013 (支系A), Yan et al., 2021 (KIZ07980+KIZ07984贵州赤水样品组成的支系, Fig. 2b, Fig. S1), Wang et al., 2021 (Q. robertingeri)。此外, Yan et al. (2013)、Yan et al. (2021) 以及Wang et al. (2021) 的数据均显示在四川合江和贵州赤水确实存在多个线粒体支系同域分布。整合Wang et al. (2021) 的数据以及更多未发表COI基因片段数据, 我们重新分析发现, Yan et al. (2013) 的Cytb支系A和支系B-D分别对应Wang et al. (2021) 的合江棘蛙和棘腹蛙COI支系。我们推测:不同基因片段的进化速率差异是导致不同研究出现树形差别的原因, 相比较而言, Wang et al. (2021) 所使用的COI基因片段具有最快的进化速率, 不适合用于解决物种之间的关系。此外外群的选择也可能是造成目前树形差异的一个因素(外群的选择应该遵循最近姐妹群原则)。最后, 综合Yan et al. (2013) 和Yan et al. (2021) 研究中基于核基因数据揭示的合江棘蛙地模群体和棘腹蛙各支系间存在基因交流, 现阶段我们仍然建议将合江棘蛙作为棘腹蛙的同物异名 (Che et al., 2009) 处理。但是我们认识到, 对于这两物种的关系需要进一步的研究确定。Yan et al. (2013) 和Yan et al. (2021) 中使用的核基因样本 (在线粒体群体数据基础上进行抽样) 及核基因信息量均较少。因此, 建议未来研究中: 在种群样本的基础上, 开展形态特征量化分析, 利用新一代的测序技术, 如简化基因组数据开展近缘种及种群水平的谱系地理学研究, 在样本量和基因数据量均增加的基础上, 相信我们会对合江棘蛙与棘腹蛙以及与其他近缘物种的关系、物种有效性等问题有更清晰的认识。随着研究的不断深入, 发现线粒体支系的替代现象在包括两栖动物在内的不同类群中较为普遍, 因此在使用单一线粒体基因片段进行物种快速鉴定的时候一定要非常小心。开展相关研究时, 总体上, 鼓励在种群水平上, 考虑生态因素, 开展量化细致的形态数据、线粒体以及核基因数据的比较整合分析。编写: 张栋儒审核: 于中斌编辑: 张栋儒2021.09.10
- 2023-10-27 张栋儒修改了地理分布
- 2023-04-25 张栋儒修改了地理分布
- 2022-03-17 张栋儒修改了讨论
- 2022-03-17 张栋儒修改了讨论
- 2021-11-26 张栋儒修改了讨论
- 2021-09-16 张栋儒修改了讨论
- 2021-09-13 张栋儒修改了讨论
- 2021-09-09 张栋儒修改了讨论
- 2021-09-03 张栋儒修改了讨论
- 2021-09-03 张栋儒修改了讨论
- 2021-09-03 张栋儒修改了讨论
- 2021-09-03 张栋儒修改了讨论
- 2020-07-08 沈文菁修改了地理分布
- 2020-07-08 沈文菁修改了蝌蚪
- 2020-07-08 沈文菁修改了卵
- 2020-07-08 沈文菁修改了皮肤、体色
- 2020-06-27 陈宏满修改了皮肤、体色
- 2020-06-27 陈宏满修改了蝌蚪
- 2020-06-27 陈宏满修改了卵
- 2020-06-27 陈宏满修改了讨论
- 2015-04-23 张宝林添加此条目
451
主要分类系统
-
中国两栖类 (2026)
-
无尾目 Anura
-
叉舌蛙科 Dicroglossidae
-
棘胸蛙属 Quasipaa
-
棘腹蛙 Quasipaa boulengeri
-
Amphibian Species of the World (2026)
-
无尾目 Anura
-
叉舌蛙科 Dicroglossidae
-
棘胸蛙属 Quasipaa
-
棘腹蛙 Quasipaa boulengeri
-
AmphibiaWeb (2026)
-
无尾目 Anura
-
叉舌蛙科 Dicroglossidae
-
棘胸蛙属 Quasipaa
-
棘腹蛙 Quasipaa boulengeri
-
中国两栖动物及其分布彩色图鉴 (2012)
-
无尾目 Anura
-
叉舌蛙科 Dicroglossidae
-
棘胸蛙属 Quasipaa
-
合江棘蛙 Quasipaa robertingeri






