蛙属
编写:张栋儒
审核:卢宸祺
编辑:张栋儒
2022.01.09
蛙属的分类近年来经历了较大变动,目前仍存在很大的争议。形态特征同型性(morphological homoplasy)(非同源性状)长期以来给蛙属的分类学带来了困难。例如,费梁等, 1990依据威宁蛙,梭德氏蛙,越南趾沟蛙Rana johnsi和桑植蛙等物种的成体趾端膨大呈小吸盘,有腹侧沟等主要性状与其它林蛙 brown frogs 类群等物种有明显区别,而将上述4个种(亚种)另立为新属,趾沟蛙属Pseudorana,随后,趾沟蛙属物种组成经历了多次的变动(例如 Dubois, 1992; 江建平等, 1997; 费梁等,2012)。建立在梭德氏蛙的蝌蚪体腹面具有大的吸盘,费梁等, 2000 建立新属拟湍蛙属 Pseudoamolops。
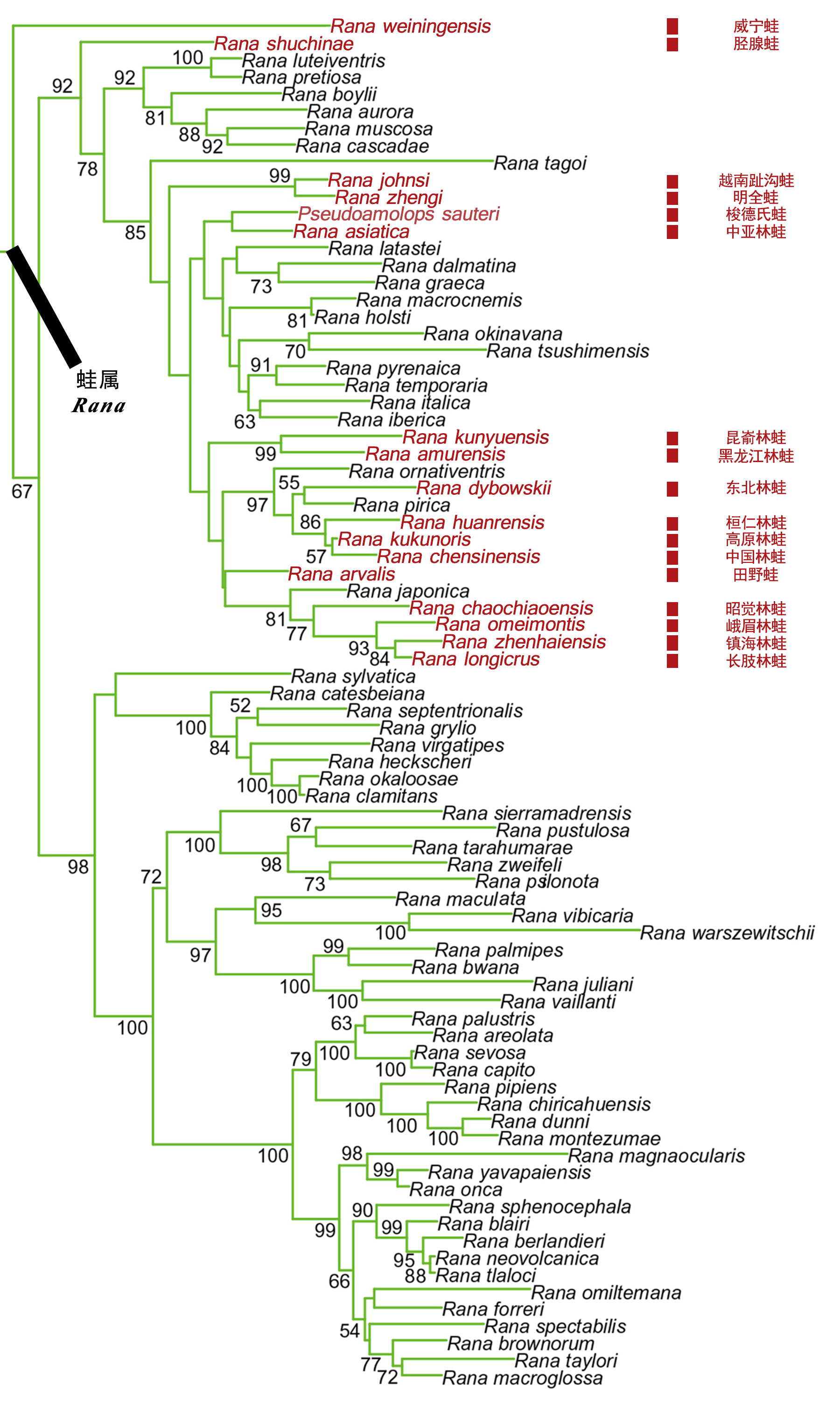
目前分子系统学研究(例如 Tanaka-Ueno et al., 1998; Che et al., 2007; Yuan et al., 2016)已经表明,上述所使用的形态性状在蛙科物种中经历了多次独立进化产生,因此,这些非同源性状不适合作为独立特征进行属级评定,拟湍蛙属和趾沟蛙属不应该成立。非常明显地,建立在Che et al., 2007;Pyron and Wiens, 2011: Fig. 2; Yuan et al., 2016: Fig.1系统发育树来看,所谓“趾沟蛙属”物种没有聚在一起呈现单系。威宁蛙位于整个蛙属的最基部;越南趾沟蛙和明全蛙聚为单系,该支系与欧亚大陆其它林蛙群物种聚成的单系互为姐妹支;梭德氏蛙与东亚内陆及岛屿其它林蛙群物种聚在一起,尽管系统演化地位没有得到明确解决(Yuan et al., 2016)。中国分布的蛙属物种与亚洲岛屿地区、欧洲及美洲等多个国家地区的物种有较为密切的关系,中国分布物种不是一个单系(Yuan et al., 2016)。
Frost et al., 2006建立在分子系统学研究基础上,将美洲部分物种建立Lithobates属,同时,该研究认为趾沟蛙属为蛙属的同物异名,因而趾沟蛙属无效。后期,建立在ASW, https://amphibiansoftheworld.amnh.org/又重新恢复了趾沟蛙属(包括中国的威宁蛙和桑植蛙),并将相关物种划分为三个属,包括趾沟蛙属(Pseudorana)、蛙属(Rana)和Lithobates属,然而根据本团队未发表的数据(车静个人通讯),威宁蛙和桑植蛙不呈现单系,从而否定了趾沟蛙属的划分。尽管分布在美洲的大部分物种在形态上相较欧亚大陆物种,具有较大形态分化,然而,例如美洲的R. sylvatica仍然保持了和欧亚林蛙类群一致的形态特征,如颞部具三角形黑斑等特征,被长期认为是典型的林蛙群物种(Che et al., 2007)。
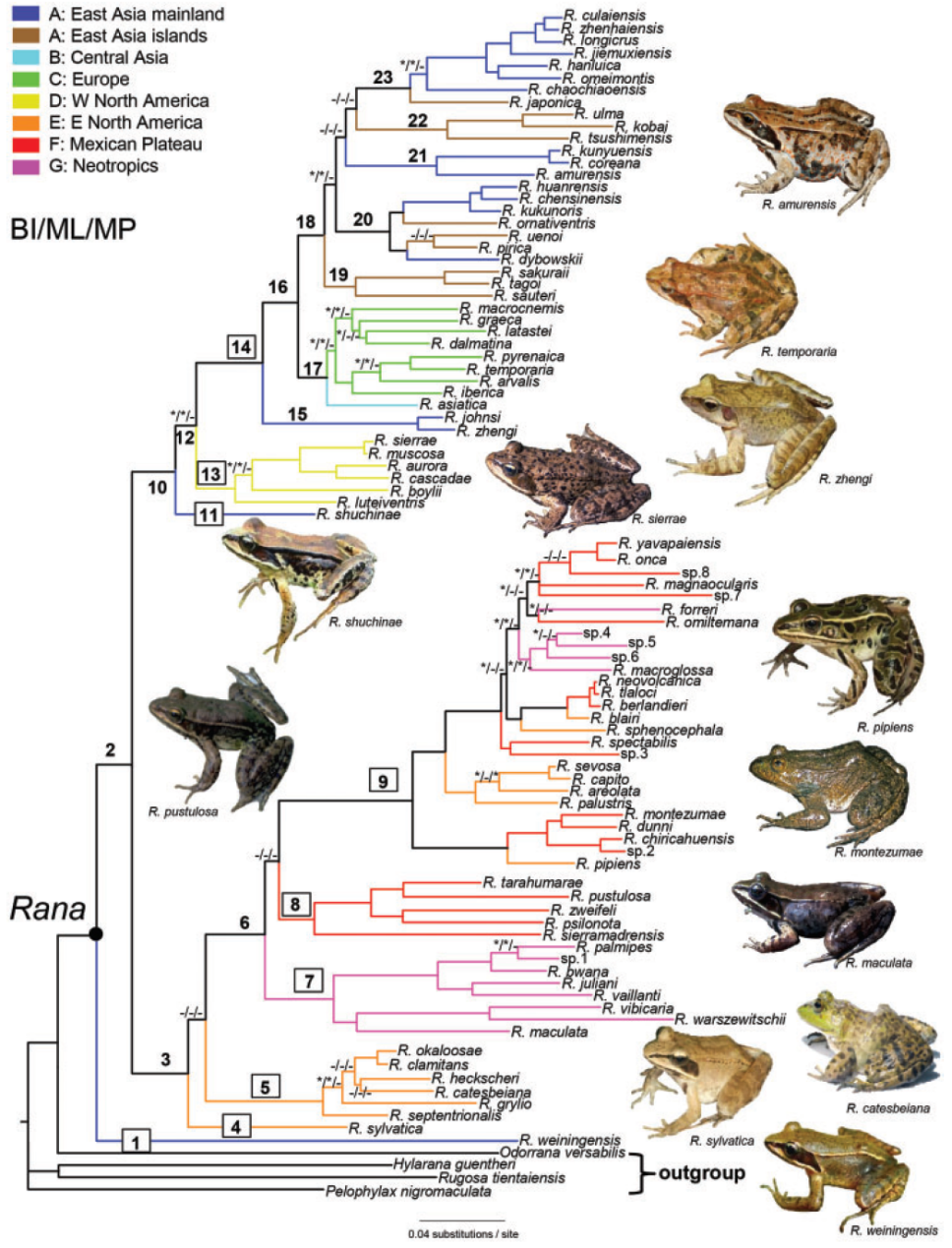
Yuan et al., 2016对蛙属整个分布区的物种进行了分子系统学研究,结果显示分布于中国西南地区的威宁蛙位于整个蛙属的最基部;其余物种分为两支,一支由西北美R. boylii种组与除威宁蛙之外的欧亚大陆蛙属物种组成,其余美洲的物种聚为另一支,因此美洲的两支蛙属物种呈现出并系结构;欧洲与中亚地区的蛙属物种聚为一支。此外,Yuan et al., 2016也整体阐释了蛙属物种在欧亚大陆和美洲大陆的演化问题:蛙属物种起源于东亚,并且分别经历了两次从东亚到新大陆地区东北美和西北美的扩散事件,以及一次从东亚进入欧洲和中亚地区的扩散事件。
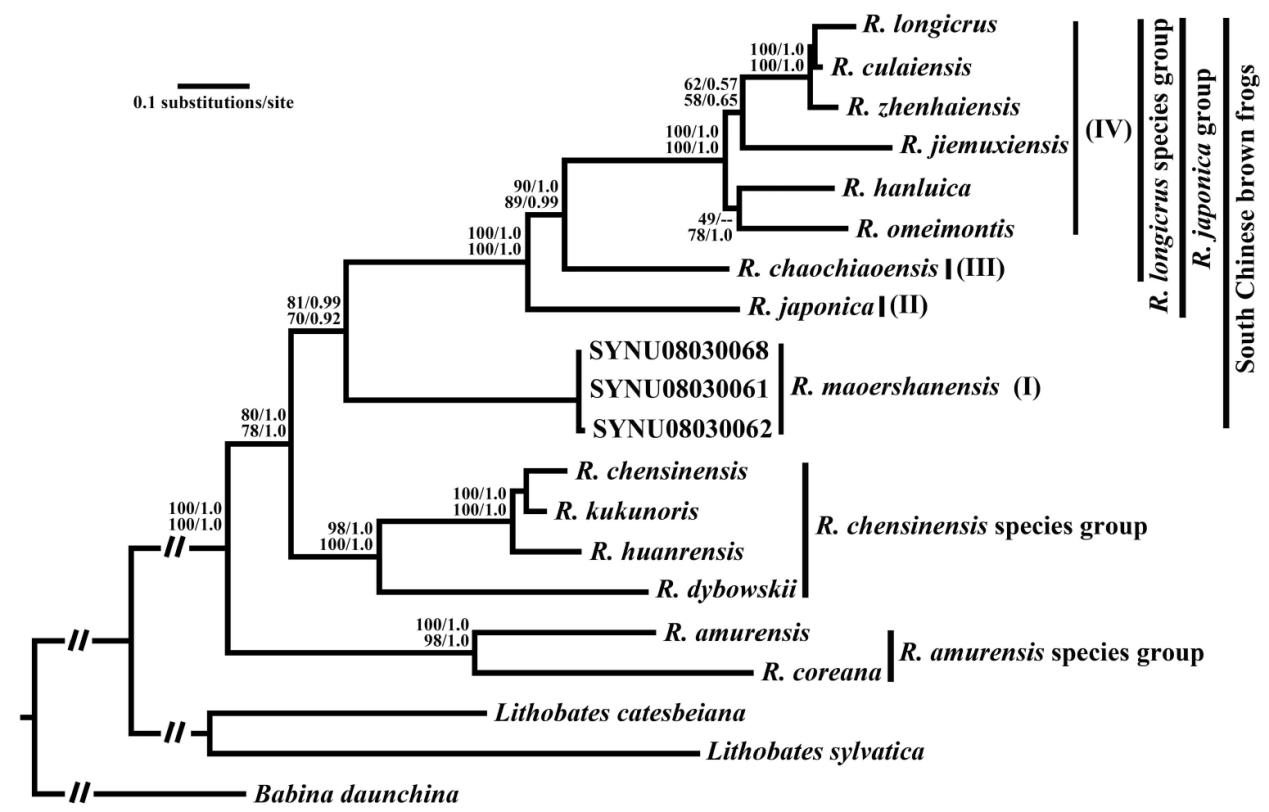
Zhou et al., 2016对东亚内陆及岛屿的林蛙群物种进行了分子系统学研究,日本林蛙种组(长肢林蛙、徂崃林蛙、镇海林蛙、借母溪林蛙、寒露林蛙、峨眉林蛙、昭觉林蛙、日本林蛙)与猫儿山林蛙互为姐妹支,这两个种组形成的单系支(中国南方林蛙群)又与中国林蛙种组(中国林蛙、高原林蛙、桓仁林蛙、东北林蛙)互为姐妹支,黑龙江林蛙种组(黑龙江林蛙、韩国林蛙)则位于最基部;在日本林蛙种组内部,日本林蛙位于最基部,昭觉林蛙与其余日本林蛙种组物种互为姐妹群。
中国在该蛙属中包括如下物种。
黑龙江林蛙 Rana amurensis
田野蛙 Rana arvalis
中亚林蛙 Rana asiatica
昭觉林蛙 Rana chaochiaoensis
中国林蛙 Rana chensinensis
峰斑林蛙 Rana chevronta
韩国林蛙 Rana coreana
徂徕林蛙 Rana culaiensis
大别山林蛙 Rana dabieshanensis
东北林蛙 Rana dybowskii
寒露林蛙 Rana hanluica
桓仁林蛙 Rana huanrensis
借母溪林蛙 Rana jiemuxiensis
九岭山林蛙 Rana jiulingensis
越南趾沟蛙 Rana johnsi
高原林蛙 Rana kukunoris
长肢林蛙 Rana longicrus
栾川林蛙 Rana luanchuanensis
猫儿山林蛙 Rana maoershanensis
姬林蛙 Rana minuscula
峨眉林蛙 Rana omeimontis
淮源林蛙 Rana qui
桑植蛙 Rana sangzhiensis
梭德氏蛙 Rana sauteri
胫腺蛙 Rana shuchinae
太行林蛙 Rana taihangensis
威宁蛙 Rana weiningensis
武夷林蛙 Rana wuyiensis
镇海林蛙 Rana zhenhaiensis
织金林蛙 Rana zhijinensis
田野蛙 Rana arvalis
中亚林蛙 Rana asiatica
昭觉林蛙 Rana chaochiaoensis
中国林蛙 Rana chensinensis
峰斑林蛙 Rana chevronta
韩国林蛙 Rana coreana
徂徕林蛙 Rana culaiensis
大别山林蛙 Rana dabieshanensis
东北林蛙 Rana dybowskii
寒露林蛙 Rana hanluica
桓仁林蛙 Rana huanrensis
借母溪林蛙 Rana jiemuxiensis
九岭山林蛙 Rana jiulingensis
越南趾沟蛙 Rana johnsi
高原林蛙 Rana kukunoris
长肢林蛙 Rana longicrus
栾川林蛙 Rana luanchuanensis
猫儿山林蛙 Rana maoershanensis
姬林蛙 Rana minuscula
峨眉林蛙 Rana omeimontis
淮源林蛙 Rana qui
桑植蛙 Rana sangzhiensis
梭德氏蛙 Rana sauteri
胫腺蛙 Rana shuchinae
太行林蛙 Rana taihangensis
威宁蛙 Rana weiningensis
武夷林蛙 Rana wuyiensis
镇海林蛙 Rana zhenhaiensis
织金林蛙 Rana zhijinensis