蜥蜴动态视觉信号
作者:齐银
1. 动态视觉信号定义
与声音和颜色信号不同,许多动物采用动态视觉信号进行通讯。如图1:婆罗洲岩蛙(Starois pavus)后肢摆动行为,西部鸊鷉(Aechmophorus occidentalis)婚舞行为和招潮蟹(Uca perplexa)螯足挥舞行为。动态视觉信号(visual motion signal)主要通过动物肢体在空间的速度和方向变化来传递信息,是一种三维立体信号。这种信号的接受主要通过视网膜边缘的神经节细胞来完成。采用动态视觉信号进行通讯的物种,其视觉系统往往具有两个视觉小凹,中间的视觉小凹负责提取动态信号的深度信息,远端的视觉小凹负责提取动态信号的平面信息。经过双视觉小凹对信号进行整合之后,传递到大脑。相比于声音和颜色信号,关于动物动态视觉信号的功能和进化的研究才刚刚起步(Stevens, 2013)。
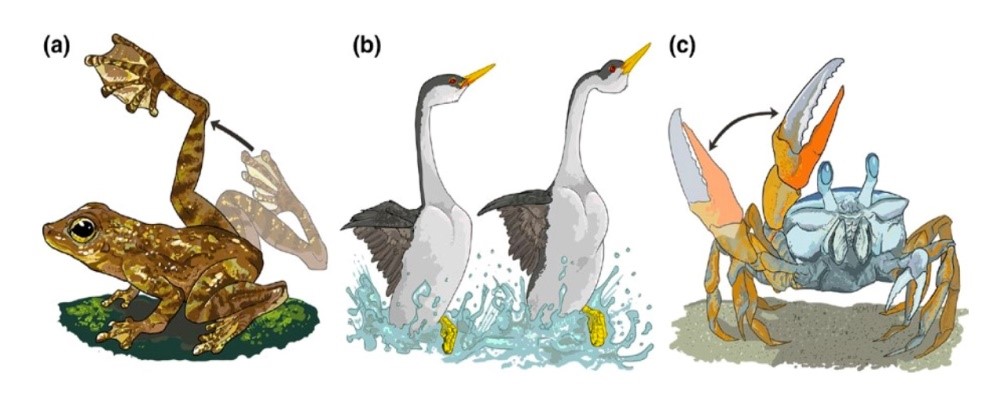
图1 (a) 婆罗洲岩蛙后肢摆动行为; (b) 西部鸊鷉婚舞行为; (c) 招潮蟹螯足挥舞行为。此图摘自(Schuppe and Fuxjager, 2017)
2. 蜥蜴动态视觉信号及其多样性
在爬行动物蜥蜴亚目(Lacertilia)中,鬣蜥科(Agamidae)、避役科(Chamaeleonidae)和美洲鬣蜥超科(Iguanas)大多数物种以及石龙子科(Scincidae)少数物种采用动态视觉信号进行通讯。蜥蜴动态视觉信号成分非常多样,包括:躬背、摆臂、身体膨大、身体抬升、身体倾侧、体色改变、冠鳞竖起、尾部展示、喉部展示和舌部展示等,而不同物种动态视觉信号往往由不同信号成分构成,且具有特定的编排次序。例如:澳洲龙蜥 (Amphibolurus muricatus)动态视觉信号由弹尾(tail-flick)、后摆臂(backward arm-wave)、前摆臂(forward arm-wave)、俯卧撑(push-up)和身体晃动(body-rock)构成(图2)(Peters et al., 2002),而对于广泛分布于亚洲和阿拉伯地区的沙蜥属物种(Phrynocephalus)而言,其动态视觉信号主要由卷尾(tail coil)和甩尾(tail wave)构成(图3),部分物种如大耳沙蜥(P. mystaceus)还表现出身体倾侧和耳部褶皱膨大行为,而荒漠沙蜥(P. przewalskii)还表现出身体旋转(body circling)和四肢拍打地面(arm drum)行为(图4)。

图2 澳洲龙蜥(Amphibolurus muricatus)动态视觉信号
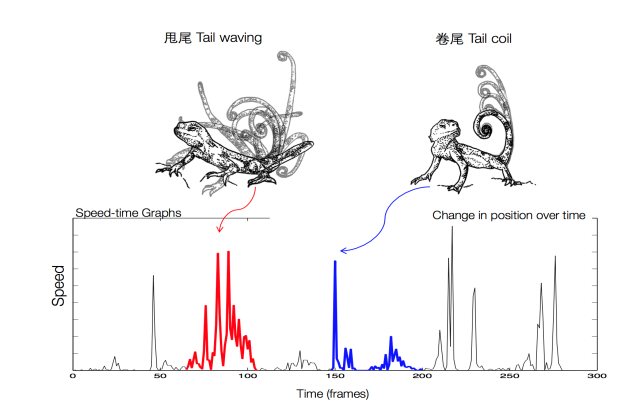
图3 沙蜥动态视觉信号基本结构

图4 大耳沙蜥和荒漠沙蜥动态视觉信号
3. 蜥蜴动态视觉信号功能
已有研究表明,蜥蜴动态视觉信号主要包括三个功能:1)作为颜色信号的辅助手段,以提升颜色信号的传递效率(Mcelroy et al., 2007);2)作为独立的信号,向对手传递特定的信息,例如个体的耐力、战斗能力和社会地位等 (Perry et al., 2004; Qi et al., 2011; Mowles and Ord, 2012)。动态视觉信号的产生涉及全身各个器官系统的协作,其信号的精准性对大脑、肌肉和身体协调性都有很高的要求,只有生理状态较好的个体,才能做出复杂的动态视觉信号 (Schuppe and Fuxjager, 2017);3)作为独立信号的前奏,以吸引信号接收者对后续信号的关注,这方面的功能主要是为应对特定的环境条件,例如较暗的光线和较高的植被盖度等。澳洲龙蜥(A.muricatus) 雄性弹尾行为可以吸引其他个体对摆臂、俯卧撑和身体晃动行为的关注,而在风速较大的情况下,个体会通过延长弹尾时间来确保吸引的效果 (Peters and Evans, 2003; Peters et al., 2007)。
4. 蜥蜴动态视觉信号适应性及其进化
由于动态视觉信号运动的特点,环境中任何一种风吹草动都会对动态视觉信号的传递和接收造成干扰,为了确保信号传递的有效性,采用动态视觉信号通讯的蜥蜴物种会面临信号复杂性调整的压力(Fleishman, 1988; Endler, 1992)。动态视觉信号复杂性可以通过增加信号成分来调整。例如:澳洲龙蜥(A.muricatus)守卫领域时,会向对手展示许多肢体动作,包括尾尖摆动、右前肢逆时针绕环、右前肢顺时针绕环、俯卧撑和背弯曲(Peters and Evans, 2003)。但是,由于动态视觉信号表达需要机体不同器官的协作,是一种高耗能过程,因此单独依靠信号成分增加来提高信号复杂性的方式易受能量代谢的限制(Matsumasa and Murai, 2005)。在这种情况下,动物更多的是通过增加修饰物或提高信号成分的内在变化,来提高动态视觉信号的复杂性(Peters et al., 2007; Ord and Stampsb, 2008)。
目前,关于蜥蜴动态视觉信号复杂性的进化,主要有两个假说:(1)中性假说(neutral hypothesis),该假说认为动态视觉信号复杂性的进化是一个中性的过程,主要受随机漂变的影响,因此动态视觉信号复杂性进化与物种演化的历史相同(Freeberg et al., 2012; Ord and Garcia-Porta, 2012);(2)适应性假说(adaptation hypothesis):该假说认为动态视觉信号复杂性进化是适应的结果,其进化方向受各种选择压力的影响(Freeberg et al., 2012; Ord and Garcia-Porta, 2012)。根据选择压力的差异,适应性假说又可分为三种:① 功能适应性(function adaptation):即动态视觉信号复杂性的增加主要基于信号功能的增加,不同信号成分表示不同的生物信息(multiple message)或相同信息的不同方面(redundancy),从而有助于信号接收者更准确的判断对手和进行物种识别(Hebets and Papaj, 2005; Hebets, 2011);② 效率适应性(efficacy adaptation):即动态视觉信号复杂性的增加主要基于信号传递和感知效率的提高,信号成分之间相互影响、相互促进,从而确保信号的有效传递(Hebets and Papaj, 2005; Hebets, 2011);③ 社会适应性(sociality adaptation):即动态视觉信号复杂性的增加主要基于社会复杂性的提高,不同的社会交流需要不同的信号成分,从而有助于维持社会秩序和避免社会冲突。受经典性选择理论的影响,许多关于动态视觉信号复杂性进化的研究都围绕功能适应性假说,即动态视觉信号复杂性的增加是否有利于配偶识别和竞争(Barske et al., 2011; Girard et al., 2015; Echeverri et al., 2017),而关于中性假说和其它适应性的研究还非常少。
沙蜥属物种是研究动态视觉信号功能和进化的良好模型
沙蜥属包括31个物种,是鬣蜥科中已知的唯一全部采用动态视觉信号进行通讯的动物类群。相比于美洲的安乐蜥属(Anolis),沙蜥属分布范围更广,从阿拉伯半岛往东经伊朗高原、中亚五国、印度、尼泊尔、俄罗斯、中国和蒙古都有分布(图5),分布生境涉及荒漠、戈壁和荒漠草原。此外,该属物种密度普遍较高,而且动态视觉信号复杂性在不同物种间差异明显,有些物种信号成分非常复杂,而有些物种则相对简单,这为动态视觉信号量化、信号结构进化历史重建和采用行为生态学方法进行信号功能研究提供了良好条件。更为重要的是,雌性沙蜥也具有非常复杂的动态视觉信号,这与美洲安乐蜥和澳洲龙蜥形成明显对比,从而为研究动态视觉信号的两性异型和雌性动态视觉信号的进化提供了良好模型。
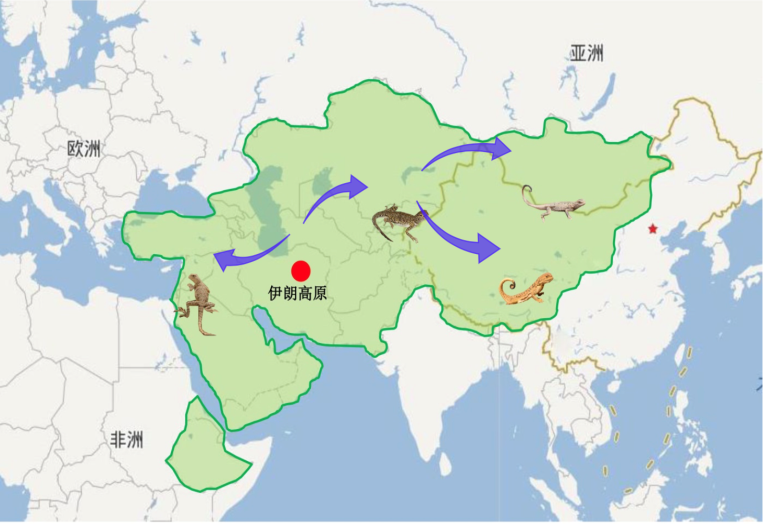
图 5 全球沙蜥分布图
参考资料
Barske, J., Schlinger, B.A., Wikelski, M., Fusani, L., 2011. Female choice for male motor skills. Proceedings of the Royal Society B: Biological Sciences 278, 3523-3528.
Echeverri, S.A., Morehouse, N.I., Zurek, D.B., 2017. Control of signaling alignment during the dynamic courtship display of a jumping spider. Behav. Ecol. 28, 1445-1453.
Endler, J., 1992. Signals, signal conditions, and the direction of evolution. The American Naturalist 139, S125-S153.
Fleishman, L.J., 1988. Sensory and environmentailn fluences on display form in Anolis auratus, a grass anole from Panama. Behav. Ecol. Sociobiol. 22, 309-316.
Freeberg, T.M., Dunbar, R.I.M., Ord, T.J., 2012. Social complexity as a proximate and ultimate factor in communicative complexity. Philosophical Transactions of the Royal Society B: Biological Sciences 2012, 1785-1801.
Girard, M.B., Elias, D.O., Kasumovic, M.M., 2015. Female preference for multi-modal courtship: multiple signals are important for male mating success in peacock spiders. Proceedings of the Royal Society B-Biological Sciences 282.
Hebets, E., 2011. Current Status and Future Directions of Research in Complex Signaling. Current Zoology 57, I-V.
Hebets, E., Papaj, D., 2005. Complex signal function: developing a framework of testable hypotheses. Behav. Ecol. Sociobiol. 57, 197-214.
Matsumasa, M., Murai, M., 2005. Changes in blood glucose and lactate levels of male fiddler crabs: effects of aggression and claw waving. Anim. Behav. 69, 569-577.
Mcelroy, E.J., Marien, C., Meyers, J.J., Irschick, D.J., 2007. Do Displays Send Information about Ornament Structure and Male Quality in the Ornate Tree Lizard, Urosaurus ornatus? Ethology 113, 1113–1122.
Mowles, S.L., Ord, T.J., 2012. Repetitive signals and mate choice: insights from contest theory. Anim. Behav. 84, 295-304.
Ord, T.J., Garcia-Porta, J., 2012. Is sociality required for the evolution of communicative complexity? Evidence weighed against alternative hypotheses in diverse taxonomic groups. Philosophical Transactions of the Royal Society B: Biological Sciences 2012, 1811-1828.
Ord, T.J., Stampsb, J.A., 2008. Alert signals enhance animal communication in"noisy" environments. Proceedings of the National Academy of Sciences 105, 18830-18835.
Perry, G., Levering, K., Girard, I., Garland, T., 2004. Locomotor performance and social dominance in male Anolis cristatellus. Anim. Behav. 67, 37-47.
Peters, R.A., Clifford, C.W.G., Evans, C.S., 2002. Measuring the structure of dynamic visual signals. Anim. Behav. 64, 131-146.
Peters, R.A., Evans, C.S., 2003. Introductory tail-flick of the Jacky dragon visual display: signal efficacy depends upon duration. J. Exp. Biol. 206, 4293.
Peters, R.A., Hemmi, J.M., Zeil, J., 2007. Signaling against the wind: modifying motion-signal structure in response to increased noise. Curr. Biol. 17, 1231-1234.
Qi, Y., Wan, H.F., Gu, H.J., Wang, Y., 2011. Do displays and badges function in establishing the social structure of male toad-headed lizards, Phrynocephalus vlangalii? Journal of Ethology 29, 381-387.
Schuppe, E.R., Fuxjager, M.J., 2017. High- speed displays encoding motor skill trigger elevated territorial aggression in downy woodpeckers. Funtional Ecology.
Stevens, M., 2013. Sensory ecology,behaviour, & evolution. Oxford University Press, UK.
作者介绍
齐银,博士,中国科学院成都生物研究所副研究员,硕导。主要研究方向:动物行为学及适应性进化。email:qiyin@cib.ac.cn
编辑:陈宏满、高伟






